
Genética molecular
Genética molecular. Es el campo de la biología que estudia
la estructura y la función de los genes a nivel molecular empleando los métodos
obtenidos de la genética y de la biología molecular. Es utilizada en la
clasificación científica de los organismos, para determinar los patrones de
descendencia, y entre sus aplicaciones está la terapia génica. Todo esto
obtenido de la información molecular de los genes, molécula que declara sus
límites diferenciándola de la biología molecular.
Características
Gen
Un gen es la unidad física y funcional de la herencia, que
se pasa de padres a hijos. Los genes están compuestos por ADN y la mayoría de
ellos contiene la información para elaborar una proteína específica. Cada gen
tiene una localización específica en un determinado cromosoma, y el conjunto de
todos los genes, contenidos en todos los cromosomas, constituye el genoma.

Los cromosomas están constituidos por ADN (ácido
desoxirribonucleico), que codifica la información hereditaria, y por proteínas
istónicas y no histónicas. Cada cromosoma está formado por una única molécula
de ADN, en la que cada gen ocupa un segmento.
El ADN está constituido por la asociación de moléculas
llamadas nucleótidos, formadas por la unión de una molécula de fosfato, una del
azúcar desoxirribosa y una base nitrogenada. Ya que cuatro bases distintas,
adenina, guanina, timina y citosina participan en la formación de los
nucleótidos, hay cuatro tipos distintos de estos. Para formar ADN, los
nucleótidos se vinculan por sus grupos fosfato y conforman una larga hebra,
cuyas bases nitrogenadas se unen por uniones débiles pero muy específicas con
las de otra hebra. Se forman así pares de bases, que determinan que ambas
hebras, apareadas, se enrollen para dar lugar a la estructura de doble hélice.
Replicación del ADN
Es el proceso según el cual la molécula de ADN de doble hélice origina a otras dos moléculas de ADN con la misma secuencia de bases.
Esta duplicación del material genético es semiconservativa debido a que las dos cadenas complementarias del ADN parental, al separarse, sirven de molde a su vez para la síntesis de una nueva cadena, complementaria de la cadena molde, de forma que cada nueva doble hélice contiene una de las cadenas del ADN parental.
La replicación se produce durante la interfase, en el
período S o de síntesis de ADN.
Agricultura y ganadería:
Aunque pueda parecer sorprendente la genética molecular han contribuido eficazmente a las herramientas de trabajo en el uso del campo y animal. Esto se debe que debido a los conocimientos que derivan de la genética molecular podemos incrementar la calidad tanto de animales como plantas. ¿Cómo? Gracias al cruce de análisis genéticos a través del cual sabemos los genes más relevantes para el desarrollo de las características que queramos lograr.
Estos son algunos de los ámbitos en los que puede intervenir la genética molecular, aunque debes de saber qué existen más ámbitos en los que proporciona gran ayuda. Por ejemplo y posiblemente te suene, la ingeniería genética, que se trata de aquella ciencia por la que podemos modificar el material genético de los seres vivos. Muchas veces se utilizan animales para conocer el funcionamiento de enfermedades que afecten a seres humanos.
 |
| Estructura de ADN |
Bibliografía:
https://www.ecured.cu/Gen%C3%A9tica_molecular
https://geneticabioterio.wordpress.com/genetica-molecular/
https://hn.euroinnova.edu.es/blog/que-estudia-la-genetica-molecular
Genética Clásica
La genética clásica es aquella que se aproxima a la genética en la cual no se emplean herramientas de biología molecular. Los primeros estudios en el campo de la transmisión de los caracteres, de la herencia genética por tanto, corresponden a este campo: por ejemplo, las Leyes de Mendel o el análisis del ligamiento. Pese al advenimiento de la genética molecular, los enfoques de la clásica siguen siendo utilizados: por ejemplo, en el mundo de la agricultura, durante la mejora genética; o, en la cartografía genética de baja resolución, a fin de situar la posición relativa de los genes en los cromosomas.
Algunas veces se emplea el término genética clásica como un sinónimo sensu lato de genética directa, oponiéndola no a la genética molecular sino a la genética inversa. Ambas difieren en el enfoque empleado durante el diseño de experimentos. De este modo, la genética directa busca situar y clonar un determinado gen de interés partiendo de individuos cuyo fenotipo es llamativo (generalmente mutantes), mientras que la genética inversa parte del conocimiento de la secuencia de ADN de los genes putativos y emplea herramientas de genética molecular y de transgénesis para generar mutantes que diluciden la función del gen.
Las leyes de Mendel.
Los descubrimientos de Mendel pueden resumirse en tres leyes, que constituyen los fundamentos básicos de transmisión genética. La terminología que empleo Mendel es de difícil comprensión; por ello vamos a utilizar la terminología actual.
Primera ley de Mendel.
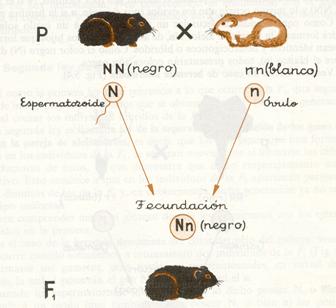
Llamada también ley de la uniformidad de los híbridos de la primera generación, dice que: cuando se realiza el cruzamiento entre dos individuos de la misma especie pertenecientes a dos variedades o razas puras (homocigóticos) todos los híbridos de la primera generación filial son iguales.
En la actualidad esta ley expresa así. “El cruce de dos razas puras da una descendencia híbrida uniforme tanto fenotípica como genotípicamente.”
Esta uniformidad de todos los individuos de la F1 puede manifestarse, bien por parecerse a uno de los padres (herencia dominante), bien porque aparezca un fenotipo con aspecto intermedio (herencia intermedia). Veamos seguidamente un ejemplo de cada caso.
Empecemos por la herencia dominante
Segunda ley de Mendel
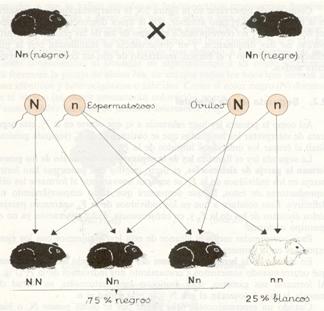
Así como la primera ley hace referencia a lo que ocurre en la Fl, esta segunda trata de interpretar los resultados que se obtienen en la F2 (segunda generación filial) al cruzar los individuos híbridos de la Fl.
La segunda ley es llamada ley de la separación o disyunción de los genes que forman la pareja de alelomorfos, es decir, que los dos genes que han formado pareja en los individuos de la Fl, se separan nuevamente al formarse las células reproductoras de éstos, lo que demuestra que dicho emparejamiento no es definitivo. Esto conduce a que en los individuos de la F2 aparezcan parejas de alelos distintos de los de la Fly, en consecuencia, dicha generación ya no es de genotipo uniforme.
Así, puede formularse esta ley actualmente: “Al cruzar entre sí los híbridos obtenidos en la primera generación, los caracteres antagónicos que poseen se separan y se reparten entre los distintos gametos, apareciendo así varios fenotipos en la descendencia”
Para comprender mejor el alcance de esta ley, seguiremos con los ejemplos expuestos en la primera.
En el caso de la herencia dominante del color del pelo del cobaya, veamos qué ocurre cuando sometemos a cruzamiento dos individuos de la Fl. Al formarse sus gametos, sean óvulos o espermatozoides, en virtud de la meiosis, la mitad poseerán el gen N y la otra mitad el n.
Tercera ley de Mendel.
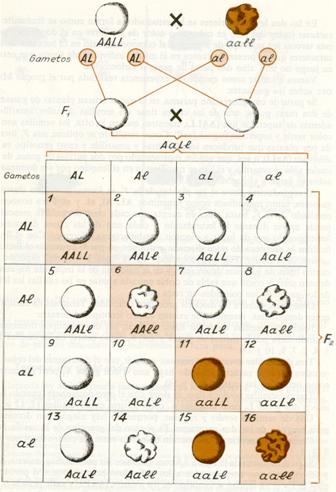
Llamada ley de la herencia independiente de los caracteres, porque expresa el hecho de que cada uno de los caracteres hereditarios se transmite a la descendencia con absoluta independencia de los demás.
Hoy se enuncia esta ley así: “Los distintos caracteres no antagónicos se heredan independientemente unos de otros, combinándose al azar en la descendencia”
En las dos leyes anteriores se ha estudiado la forma como se transmite un carácter (color del pelo en cobayas o color de las flores en el dondiego); pero esta tercera ley se ocupa de averiguar el comportamiento en la herencia de dos caracteres que se presentan juntos en el mismo individuo, de suerte que entran en juego no uno, sino dos pares de genes o alelomorfos (dihibridismo ).
Vamos a tomar como ejemplo la experiencia realizada por el propio Mendel sobre los guisantes:
Bibliografía:
https://es.wikipedia.org/wiki/Gen%C3%A9tica_cl%C3%A1sica
https://www.biologiasur.org/index.php/herencia/genetica-mendeliana
Genética evolutiva y de poblaciones
La genética cuantitativa es la rama de la genética que
estudia los caracteres controlados por muchos genes, denominados poligénicos,
y de sus propiedades genéticas en las
poblaciones.

Tipos de caracteres
Las observaciones que sirvieron de base a las Leyes de la Herencia se realizaron en caracteres cualitativos que expresaban diferencias de clase entre os fenotipos: individuos de color negro o marrón, pelo largo o corto, etc. Sin embargo, se define como fenotipo de un individuo a toda característica visible o medible de dicho individuo. Muchos caracteres de interés económico en producción animal, específicamente los que pueden ser medidos, varían en forma continua, lo que significa que los individuos no pueden ser clasificados en clases discretas.
Básicamente se puede hablar de dos tipos de caracteres:
Cualitativos
Son aquellos que están determinados por uno o muy pocos
pares de genes, por eso también son llamados oligogénicos. Presentan una
variación cualitativa, discreta o discontinua: en una población se observan
clases de individuos según el genotipo que presenten y el mecanismo de acción
génica actuante.

Cuantitativos
Otro tipo de características no pueden ser encuadradas dentro de la una variación discreta, ya que para ellas puede existir un espectro de fenotipos que cambian imperceptiblemente de un tipo a otro. Son denominados cuantitativos, métricos o continuos, ya que son aquellos que pueden ser medidos en los individuos: peso, altura, tamaño de camada, conversión alimenticia, etc. La mayoría de ellos, en una población, presentan una distribución normal.

Hipótesis de los factores múltiples
Surgió a principios del siglo XX, fruto de una controversia
entre aquellos científicos que sostenían que la variación genotípica continua
podría explicarse en términos mendelianos, y los que no. Fue postulada por
genéticos como William Bateson y Gudny Yule. Según esta hipótesis, muchos genes
comportándose cada uno de ellos de modo mendeliano contribuirían al fenotipo de
modo acumulativo.
Esta hipótesis se basa en gran medida en los experimentos de
Hermann Nilsson-Ehle, quien utilizó el color del grano de trigo para comprobar
que el rango acumulativo de alelos en múltiples loci daba lugar al rango de
fenotipos observado en los caracteres cuantitativos.
Sus experimentos consistían en cruzar trigo de granos rojos
con otro de granos blancos. La F1 presentaba un color rosa que parecía ser
dominancia incompleta, pero carecía de las proporciones fenotípicas 3:1. En
cambio, aproximadamente 15/16 presentaban tonalidad roja y 1/16 blancos.
Examinando la F2 observó que había 4 tonalidades de rojo diferentes. Teniendo
en cuento que las proporciones se presentaban en dieciseisavos, parecía que dos
genes, con dos alelos cada uno, controlaba el fenotipo, segregando de forma
independiente de forma mendeliana.
Así pues, esta ley consta de 5 puntos principales:
- Los caracteres fenotípicos con variación continua se pueden cuantificar.
- A menudo, dos o más genes, actúan sobre el fenotipo de modo aditivo.
- Cada locus puede estar ocupado por un alelo aditivo, que contribuye con una cantidad dada al fenotipo, o por un alelo no aditivo, que no contribuye.
- La contribución de cada alelo aditivo al fenotipo es aproximadamente equivalente.
- Juntos, los alelos aditivos dan lugar a una variación fenotípica sustancial.
Heredabilidad
En un sentido amplio, indica la proporción de la variación
fenotípica que es atribuible a la variación genética. En un sentido estricto, indica la proporción
de la variación fenotípica que es atribuible a la variación genética aditiva,
es decir, expresa la confiabilidad que tiene un determinado individuo
seleccionado de originar descendencia similar a él Varía entre 0 a 100% y en función de ella, se
definirá el método o procedimiento de mejora a utilizar.
Bibliografía:
https://geneticabioterio.wordpress.com/genetica-cuantitativa/
0 Comentarios